
БНБ
"ЭБНБ" (14252)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Векторные системы для клонирования, развитие
Определение "Векторные системы для клонирования, развитие" в ЭБНБ
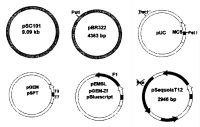 |
| Схема развития плазмидных векторов |
Так, векторы для молекулярного клонирования первого поколения представляют собой естественные бактериальные плазмиды так называемого дикого типа, обнаруженные ранее в клетках кишечной палочки Escherichia coli как экстрахромосомальные элементы. Плазмида pSC101, характеризующаяся большим размером, низкой копийностью и естественным расположением сайтов узнавания всех рестрикционных эндонуклеаз, была одним из таких векторов, который широко использовался в те первые годы молекулярного клонирования. Ее возможности как векторной молекулы были сильно ограничены и поэтому шло конструирование новых плазмид, обладающих целым рядом преимуществ.
Ко второму поколению векторов следует отнести уже искусственно созданные плазмиды, объединившие в себе черты различных природных плазмид дикого типа. Их наиболее ярким представителем явился плазмидный вектор pBR322, вобравший в себя такие черты своих предшественников, как репликон рМВ1, ген устойчивости к тетрациклину TcR и ген устойчивости к ампициллину ApS. За счет ослабленного контроля репликации копийность плазмиды pBR322 составила уже около 20-50 копий на клетку. Единственное, что осталось неизменным по отношению к первому поколению векторов, как это то же самое естественное расположение сайтов узнавания эндонуклеаз рестрикции. Плазмида pBR322 оказалась достаточно удобным вектором для молекулярного клонирования и многие ее черты легли в основу третьего поколения плазмидных векторов. Так, от pBR322 к ним перешли репликон ColEl и маркерный ген устойчивости к амплициллину. Размер самой векторной молекулы, впрочем, еще уменьшился, главным образом, за счет удаления гена устойчивости к тетрациклину. Копийность на бактериальную клетку возросла и достигла 200 копий. Однако главными новыми чертами этих плазмидных векторов можно считать маркерный ген, кодирующий NH2-концевой фрагмент β-галактозидазы LacZ' и встроенный в него искусственно созданный полилинкер, а также то, что в остальной последовательности вектора были удалены сайты узнавания некоторых рестрикционных эндонуклеаз с гексануклеотидными сайтами узнавания.
Следующее поколение плазмидных векторов, первыми представителями которого являются плазмиды pSP64 и pSP65, считать отдельным поколением несколько трудно по причине не очень значительного усовершенствования векторов предыдущего поколения, на основе которых они и были сконструированы. Однако тот факт, что практически все векторы общего назначения следующих поколений также содержат эти небольшие участки промоторных областей бактериофагов SP6, ТЗ или Т7, то их все же можно отнести к четвертому поколению.
Векторы пятого поколения, сохранив главные черты своих предшественников, весьма сильно отличаются от них, поскольку представляют собой гибридные конструкции, содержащие, кроме плазмидных последовательностей, межгенный участок одноцепочечного нитевидного фага f1, ответственный за репликацию фага. Такие химерные векторы, получившие название фагмид, содержали сразу два участка инициации репликации f1 и ColE1. Одной из первых широко применявшихся фрагмид были вектора семейства pEMBL, созданные на основе векторов pUC8/pUC9 и межгенного участка одноцепочечного фага f1.
Шестое поколение векторов, первым представителем которого является вектор pSequoiaT12, отличается от своих предшественников только тем, что в его нуклеотидной последовательности нарушены сайты узнавания для одной или нескольких частощепящих тетрануклеотидных рестрикционных эндонуклеаз. Так, первые два поколения плазмидных векторов имели естественное расположение сайтов узнавания как гексануклеотидных, так и тетрануклеотидных рестрикционных эндонуклеаз. У всех последующих поколений векторов, начиная с третьего, кроме прочих преимуществ, появился также искусственно созданный полилинкер, в котором были сосредоточены сайты узнавания нескольких гексануклеотидных рестрикционных эндонуклеаз. Однако для его эффективного использования было необходимо нарушить имевшиеся сайты узнавания этих ферментов в остальной последовательности вектора. Поэтому для векторов третьего поколения в отличие от своих предшетвенников уже не характерно естественное расположение сайтов узнавания некоторых ключевых (полилинкерных) гексануклеотидных рестрикционных эндонуклеаз в самой последовательности вектора. Однако, можно считать, что все ранее созданные векторы предыдущих поколений все же имели весьма существенный недостаток в виде наличия в их векторной последовательности сайтов узнавания для частощепящих рестрикционных эндонуклеаз с тетрануклеотидными участками узнавания, ввиду чего было невозможно расщеплять вставку этими ферментами, поскольку это неминуемо привело бы к разрушению вектора. Уникальной чертой векторов pSequoia является отсутствие в их нуклеотидной последовательности сайтов узнавания для некоторых частощепящих рестрикционных эндонуклеаз с тетрануклеотидными участками узнавания. Создание плазмидных векторов с отсутствием в них сайтов тетрануклеотидных рестрикционных эндонуклеаз значительно уступает по своей значимости созданию, например, фагмидных векторов. Однако то, что данная черта в виде отсутствия сайтов узнавания для отдельных тетрануклеотидных рестрикционных эндонуклеаз, несомненно, будет присуща последующим поколениям плазмидных векторов общего назначения, позволяет считать семейство векторов pSequoia новым, шестым поколением плазмидных векторов.
| "ЭБНБ" >> "В" >> "ВЕ" >> "ВЕК" |
Статья про "Векторные системы для клонирования, развитие" в Энциклопедии БНБ была прочитана 6512 раз
| Коптим скумбрию в коробке |
| Ананасы на гриле |
TOP 15
- Вши и борьба с ними
- Пижма обыкновенная
- Каштан конский обыкновенный
- Роль элементов в жизни растений
- Пшеница
- Иван-чай узколистный
- Обеззараживание химическими средствами
- Календула лекарственная
- Клевер луговой
- Дезинфекция текущая
- Душица обыкновенная
- Бессмертник песчаный
- Как устроены растения
- Морфологические признаки почвы
- Хвощ полевой