
БНБ
"ЭБНБ" (14252)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Тиротропин-рилизинг гормон
Определение "Тиротропин-рилизинг гормон" в ЭБНБ
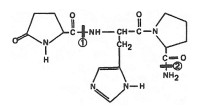 |
| Структура тиротропин-рилизинг гормона |
Предшественником ТРГ в центральной нервной системе является проТРГ, который превращается в активный гормон под действием эндопептидаз в результате ограниченного протеолиза. Синтезируется ТРГ на рибосомах клеток супраоптического и суправентрикулярного ядер гипоталамуса из макромолекулы предшественника, которая содержит небольшой участок с аминокислотной последовательностью глутамин-гистидин-пролин-глицин. После трансляции глутаминовая кислота подвергается циклизации и терминальный глицин замещается аминогруппой. Схематическое изображение последовательности путей метаболизма ТРГ представлено на рис.
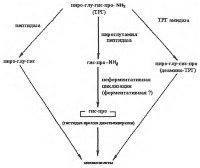 |
| Метаболизм тиротропин-рилизинг гормона |
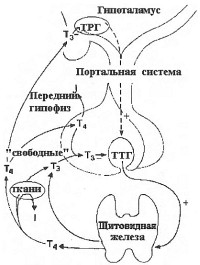 |
| Регуляция в гипоталамо-гипофизарно-тиреоидной оси |
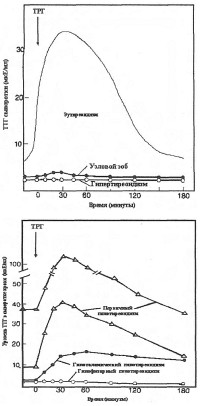 |
| Динамика тиротропин-рилизинг гормона |
ТРГ и его метаболиты обнаружены также в желудочно-кишечном тракте, островках Лангерганса поджелудочной железы, плаценте, сердце, простате, яичках и яичниках. В раннем неонатальном периоде его содержание в поджелудочной железе выше, чем в гипоталамусе, но с возрастом содержание ТРГ в гипоталамусе и мозге увеличивается, а в поджелудочной железе и желудочно-кишечном тракте уменьшается.
Предполагается, что поскольку действие ТРГ, как правило, противоположно действию соматостатина и известно, что соматостатин регулирует секрецию гормонов поджелудочной железы, то Тиротропин-рилизинг гормон может также участвовать в регуляции эндокринной функции поджелудочной железы как антагонист соматостатина. Обнаружение, подобно другим пептидам, ТРГ в ЦНС, ЖКТ, поджелудочной железе предполагает, что клетки, синтезирующие ТРГ в периферических тканях, является частью диффузной эндокринной системы организма. Синтез мРНК этого гормона в периферических тканях не ингибируется действием Т3. Роль ТРГ в этих тканях остается пока неясной.
Регуляция образования и секреции ТРГ осуществляется воздействием многих биологически активных веществ и гормонов щитовидной железы. Торможение секреции ТРГ достигается под действием соматостатина, при повышении уровня тиреоидных гормонов. Ингибирующее влияние глюкокортикоидов осуществляется в оси гипоталамус-гипофиз-щитовидная железа путем подавления ими образования ТРГ в суправентрикулярных ядрах гипоталамуса в результате прямого тормозящего действия на нейросекреторные клетки, синтезирующие Тиротропин-рилизинг гормон.
Полагают, что наблюдаемое при действии холода повышение в крови уровня тиротропина является результатом влияния на тиротрофы ТРГ, секреция которого усиливается через холодовую активацию α-адренергических структур. Подтверждением этому является возможность предупреждения роста уровня тиреотропного гормона (ТТГ введением α-адреноблокаторов до холодового воздействия. Содержание в гипоталамусе ТРГ истощается после удаления щитовидной железы у крыс и восстанавливается после ее трансплантации. Влияние гормонов щитовидной железы на секрецию ТРГ не отмечается после гипофизэктомии и в нормальных условиях эффективное воздействие тиреоидных гормонов достигается не только за счет длинной петли обратной связи (щитовидная железа - гипоталамус; тиреоидные гормоны - ТРГ), но и короткой петли обратной связи (ТТГ - ТРГ). Эти взаимодействия схематически представлены на рис.
Т3 способен непосредственно ингибировать транскрипцию гена, кодирующего синтез предшественников ТРГ и, таким образом, подавлять его синтез в гипоталамусе. Эффективным ингибитором синтеза и секреции ТРГ может быть также Т4, в случае его превращения в Т3 в пептидергических нейронах гипоталамуса.
Синтез и секреция гипоталамического ТРГ в портальную систему гипофиза регулируется уровнем тиреоидных гормонов. Секреция стимулируется при снижении в плазме крови уровней Т4 и Т3, понижении уровня Т3 в мозге, действии агонистов а-адренергических рецепторов, аргинин-вазопрессина. Секреция ТРГ угнетается при повышении уровней Т4 и Т3 в плазме крови, Т3 в мозге и действием α-адреноблокаторов.
Возбуждение как периферических, так и центральных терморецепторов модулирует синтез ТРГ в нейросекреторных клетках гипоталамуса, оказывая тем самым влияние на синтез тиреотропного гормона в гипофизе. Понижение температуры окружающей среды или температуры тела ведет к увеличению секреции ТРГ и тиротропина. Однако это влияние имеет место преимущественно у животных в эксперименте и у новорожденных детей. Влияние изменений температуры на секрецию ТРГ у взрослого человека не зарегистрировано.
ТРГ выполняет свою главную функцию - стимулирует синтез и секрецию тиротропина, после связывания с высокоаффинными рецепторами клеточной мембраны тиротрофов. Кроме того, Тиротропин-рилизинг гормон способен связываться с подобными рецепторами на мембране пролактинсекретирующих клеток и стимулировать синтез и секрецию пролактина. ТРГ рецепторы относятся к семейству трансмембранных рецепторов, связанных с G-белком. Ген, кодирующий синтез рецепторов ТРГ, расположен в 8-ой хромосоме.
Тиреоидные гормоны вызывают уменьшение числа мембранных рецепторов ТРГ тиротрофов и понижение их чувствительности к действию ТРГ. Ответ тиротрофов на действие ТРГ является двухфазным: вначале он проявляется увеличением секреции ранее синтезированного и депонированного в клетке тиреотропного гормона, а затем стимуляцией транскрипции гена и усилением синтеза тиротропина тиротрофами, поддерживающим его дальнейшую секрецию.
Связавшись с 3-им трансмембранным доменом своего рецептора на мембране тиротрофа, ТРГ передает активирующий сигнал G-белку, посредством которого активируется фермент фосфолипаза С. Фосфолипаза С гидролизует фосфатидилинозитол-4,5-дифосфат с высвобождением инозитол-1,4,5-трифосфата (ИФ3), который стимулирует освобождение внутриклеточного Са++ из его внутренних депо. Одновременно, образующийся в клетке 1,2-диацилглицерол ведет к повышению активности протеинкиназы С. Эти и ряд других внутриклеточных событий приводят к резкому повышению уровня секреции тиротропина. В более поздний период действия ТРГ (2-ая фаза) повышается вход Са++ в клетку, имеет место дальнейшая активация протеинкиназы С, активация аденилатциклазы и повышение уровня цАМФ, которые ведут к повышению экспрессии генов, кодирующих синтез альфа- и бета- субъединиц тиротропина.
Чувствительность тиротрофов к действию ТРГ регулируется дофамином и соматостатином, которые могут также тормозить секрецию тиреотропного гормона при изменении содержания тиреоидных гормонов. Секреция тиротропина и реакция тиротрофов на действие ТРГ модулируется уровнем в крови стероидных гормонов, кортизола, гормона роста. В частности, эстрогены увеличивают число мембранных рецепторов ТРГ тиротрофов и повышают их чувствительность к действию ТРГ. Возможно, что одной из функцией ТРГ является определение установочной точки реагирования или пороговой чувствительности тиротрофов к изменению содержания тиреоидных гормонов в условиях одновременного действия на тиротрофы других гормонов.
Действие ТРГ на тиротрофы имеет значение не только для активации синтеза и секреции ТТГ, но и для достижения более полного биохимического «созревания» ТТГ в результате гликозилирования полипептидных цепей α- и β- субъединиц в шероховатом эндоплазматическом ретикулуме и присоединения остатков серной (SO4) и сиаловой кислот в дистальных областях аппарата Гольджи (см. рис.). Поэтому Тиротропин-рилизинг гормон, синтезированный под действием ТРГ, обладает более высокой биологической активностью, чем тиротропин образуемый в тиротрофах при центральном гипотиреоидизме в условиях нарушения синтеза ТРГ. У больных с опухолями гипоталамуса и при гипотиреоидизме может содержаться существенное количество неактивного тиротропина.
Минимальная доза ТРГ, необходимая для стимулирования секреции ТТГ после внутривенного введения, составляет около 15 мкг. Внутривенное введение человеку 15-400 мкг ТРГ вызывает через 2-5 мин дозозависимое увеличение синтеза и секреции тиротропина гипофизом. Максимального значения концентрация ТТГ достигает в крови с 20 по 30 мин и возвращается к исходному уровню через 2-3 часа. Типичные варианты реакций на введение ТРГ приведены на рис., из которого видно, что усиленная секреция ТТГ в ответ на введение ТРГ имеет место при первичном гипотиреоидизме, а ослабленная - наблюдается при гипертиреоидизме, узловом зобе, гипофизарной форме гипотиреоидизма (вторичном гипотиреоидизме).
За увеличением уровня секреции тиротропина, вызванного введением ТРГ, следует физиологическая активация функции щитовидной железы, выражающаяся в увеличении секреции Т3 и T4, максимальное повышение сывороточного уровня которых регистрируется через 3 и 8 часов соответственно.
Дозозависимый ответ усиления гипофизом секреции ТТГ на введение ТРГ наблюдается вплоть до 400 мкг, превышение этой дозы обычно не ведет к дальнейшему увеличению секреции ТТГ. ТРГ вызывает стимуляцию секреции ТТГ и при приеме внутрь, но при этом необходимы в 2 0-40 раз большие дозы для получения того же стимулирующего секрецию ТТГ эффекта, что и при внутривенном введении. Это может быть связано, например, с частичным гидролизом пептидных связей в молекуле ТРГ в желудочно-кишечном тракте и в печени. При нормальной функции щитовидной железы чувствительность тиротрофов у женщин выше, чем у мужчин, что может быть связано с некоторой сенситизацией рецепторов ТРГ под действием эстрогенов. С возрастом эта чувствительность понижается, но у мужчин в большей степени, чем у женщин.
Тиротропин-рилизинг гормон быстро разрушается (время полужизни 2-6 мин) под действием ряда ферментов в тканях и жидкостях тела. Оказалось, что ряд его метаболитов также являются биологически активными веществами. Например, метаболит гистидилпролиндикетопиперазин проявляет большую активность как антагонист алкогольного наркоза, чем его предшественник ТРГ. В то же время этот метаболит, в противоположность ТРГ, тормозит секрецию пролактина и вызывает гипотермию. Некоторой биологической активностью обладают и другие метаболиты ТРГ. Гипоталамо-гипофизарные заболевания и, в частности, опухоли гипоталамуса и передней доли гипофиза могут приводить к разрушению секреторных клеток и сопровождаться нарушением секреции ТРГ и ТТГ. Если в эти процессы вовлечены тиротрофы гипофиза, у больного может развиться вторичный гипотиреоидизм, а если вовлечены ТРГ-нейросекреторные нейроны гипоталамуса - развивается третичный гипотиреоидизм.
| "ЭБНБ" >> "Т" >> "ТИ" |
Статья про "Тиротропин-рилизинг гормон" в Энциклопедии БНБ была прочитана 7289 раз
| Коптим скумбрию в коробке |
| Луковый соус |
TOP 15
- Вши и борьба с ними
- Пижма обыкновенная
- Каштан конский обыкновенный
- Роль элементов в жизни растений
- Пшеница
- Иван-чай узколистный
- Календула лекарственная
- Обеззараживание химическими средствами
- Клевер луговой
- Дезинфекция текущая
- Душица обыкновенная
- Бессмертник песчаный
- Как устроены растения
- Хвощ полевой
- Морфологические признаки почвы