
БНБ
"ЭБНБ" (14252)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Фораминиферы, экология и тафономия
Определение "Фораминиферы, экология и тафономия" в ЭБНБ
 |
| Породообразующие фораминиферы |
 |
| Породообразующие фораминиферы |
Вертикальное распределение планктонных фораминифер в экваториальной части Атлантики
| Глубина (в м) | Число проб | Среднее число экземпляров в пробе |
| 0-100 | 71 | 489 |
| 100-200 | 7 | 8 |
| 200-400 | 13 | 24 |
| 400-600 | 19 | 10 |
| 600-800 | 12 | 11 |
| 800-1000 | 19 | 28 |
 |
| Породообразующие фораминиферы |
 |
| Шлифы карбонатных пород |
 |
| Шлифы карбонатных пород |
 |
| Комплексы планктонных фораминифер |
 |
| Планктонные фораминиферы |
 |
| Температура воды в Карибском море |
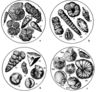 |
| Комплексы бентических фораминифер |
 |
| Колебания донной температуры |
 |
| Рост камер у Elphidium striatopunctatum |
- Литоральная зона, отличающаяся в дальневосточных морях резкими колебаниями температур (годовая амплитуда в Японском море до 20°, в Охотском 10-15° и в Беринговом 7-10°) и известным опреснением. На литорали Курильских островов были обнаружены эврифациальные Elphidium crispum (Linne) , Quinqueloculina araucana Orbigny и Q. lutea Orbigny, вряд ли представляющие собою характерные для литорали формы, поскольку встречаются и на больших глубинах.
- Сублиторальная зона (до 50 м) также отличающаяся значительными колебаниями температур. Здесь в холодноводных условиях развивается ряд видов, встречающихся в сублиторали арктических морей: Hippocrepina indiuisa Parker, Reophax curtus Cushman, Аmmоbaculites cassis (Dawson), Spiroplectammina biformis (Parker et Jones), Verneuilina advena Cushman, Trochammina fissuraperta Stschedrina, Elphidium oregonense Cushman et Grant и др. В областях влияния тихоокеанских теплых вод встречаются Quinquloculina caveriana Orbigny, Nonion boueanus (Orbigny), Streblus beccarii (Linne) и др.
- Псевдоабиссальная зона (50-200 м). Донная температура в Охотском море (по Саидовой, 1955) 1,5°, соленость менее 33,5%o. Холодноводная группа видов, характерная для северных частей дальневосточных морей и распространяющаяся к югу, следуя за холодными течениями, представлена общими и для арктических бассейнов Reophax findens (Parker), Miliammina agglutinata (Cushman), Labrospira crassimargo (Norman), Robertina arctica Orbigny и др. Специфические для дальневосточных морей формы: Alveolophragmium orbiculatum Stschedrina с его разновидностью var. ochotensis Stschedrina, Textularia conica (Orbigny) и др.
В этой зоне, в областях распространения тихоокеанских вод встречаются Robulus calcar (Linne), R. cultratus Montfort, Eponidus rераndus (Fichtel et Moll), Nonion grateloupi (Orbigny), Bulimina auriculata (Bailey).
Развитие фораминифер
Батиальная зона или область материкового склона отличается следующими условиями (по Саидовой, 1955): температура воды 0-2,4°, соленость 33,75-34,5%о.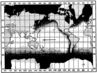
Распределение холодноводных фораминифер - Верхняя часть батиальной зоны (200-750м). Характерны общие и для арктического бассейна Rhabdammina discreta Brady, Rh. cornuta Brady, Cornuspira involvens Reuss и др. Специфические для дальневосточных морей формы: Cassidulina califotnica Cushman, Karreriella baccata (Schwager) и др.
- Нижняя часть батиальной зоны (750-2000м). Распространены встречающиеся и в дальневосточных и в арктических морях Haplophragmoides subglobosus (Sars), Dentalina obliqua (Linne), Eponides tenerus (Brady) и др. Специфические дальневосточные формы: Chilostomellina fimbriata Cushman и др.
- Абиссальная зона (2000-5000 м). Донная температура во впадинах Охотского моря 1,8-2,3º, соленость 34,5-34,7%o. Общие для Тихого и Северного Ледовитого океанов: Astrorhiza crassatina Brady, Reophax distans Brady, R. guttifer Brady и др. Тихоокеанские виды: Rhabdammina linearis Brady, Trochammina globigeriniformis (Parker et Jones) и др.
- Щедрина выделяет для Тикоокеанского бассейна еще одну зону - сверхабиссальную (свыше 5000 м), для которой характерны Reophax bacillari Brady, Hormosina globulifera Brady и др.
 |
| Рраспространение тепловодных фораминифер |
Так, для области, расположенной к югу от Флориды, Нортоном (Norton, 1930) были установлены следующие зоны, для донных осадков, каждой из которых характерны свои виды фораминифер:
- От берега до глубины 9 м, колебания температур 25,5-31,4°. Преобладают Miliolidae и Peneroplidae, местами Valvulinidae (Ataxophragmiidae), Саlcarinidae, Homotremidae и Nonionidae.
- Глубины 9-110 м, температуры 18,9-24,8°. Господствуют Lagenidae, Textulariidae, Buliminidae, Amphisteginidae; количество Miliolidae и Peneroplidae несколько сокращается.
- Глубина 900-1500 м, температура 4,0-7,6°. Преобладают планктонные Globigerinidae и Globorotaliidae; значительную роль играют Lagenidae и Buliminidae, Anomalinidae встречаются на всех глубинах. Комплекс видов смешанный; в его состав входят обитающие на дне и попавшие из пелагиали формы.
- Глубина 3650-5200 м, температура 1,8-2,0°. Комплекс видов, близкий к распространенному в зоне C.
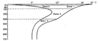
Зависимость распространения современных фэраминифер в более северном районе о. Лонг Айленд (у берегов Северной Америки) от условий температуры, глубины и отчасти солености была изучена Фледжером (Phleger, 1945) и Паркер (Parker, 1948).
В работе Паркер приведен весьма наглядный график изменения максимальных и минимальных годовых температур с глубиной и указаны изменения солености морской воды в пределах верхних 300 м (рис). Паркер выделяет следующие зоны, характеризующиеся каждая своим комплексом видов фораминифер:
- Глубина 0-15 м. Обилие Elphidium и Streblus baccarii (Linne), Quinqueloculina seminulum (Orbigny), Eggerela advena Cushman.
- Глубина 15-90 м. Преобладание Miliolidae, Eggerella advena (Cushman), Trochammina lobata, Buliminidae, Polymorphinidae и некоторые Nonionidae и Anomalinidae.
- Глубина 90-300 м. Преобладание Lagenidae, почти исключительно приуроченных к этой зоне; кроме того, присутствуют Textulariidae, Trochamminidae, Miliolidae, Discorbidae, Buliminidae, Anomalinidae, Nonionidae, Epistominidae, Chilostomellidae.
- Глубина 300-700 м. Lagenidae становятся очень редкими. Преобладают Buliminidae, Ammosphaeroidina sphaeroidiniformis (Brady), Martinottiella nodulosa (Cushman), Cassidulinita; реже встречаются Valvulina conica (Brady), Karreriella bradyi (Cushman), Pseudoglandulina occidentalis (Cushman) и Bolivina subspinescens (Cushman).
Интересные и новые данные касаются зонального распределения фораминифер в Карибском море (Lowman, 1949). Меридиональный профиль, по которому отбирались образцы, начинается в пресноводных лагунах дельты Миссисипи, пересекает район полузамкнутых опресненных заливов, область солоноватоводного мелкого моря, материковой ступени, материкового склона и заканчивается в абиссали.
Сходная с установленной для Атлантического и Карибского морей зональность была выявлена для Тихого океана у побережья Калифорнии (Natland, 1933). Здесь было установлено пять зон.
- Неглубокие воды сублиторали 0,3 м при отливе и 1,5-2 м во время прилива, температура воды в августе 24,5°, в январе 5,7°. Встречаются Trochammina inflata (Montagu) и Streblus beccarii (Linne). Последний вид у Тихоокеанского побережья Центральной Америки спускается до глубины 100 м, где температура 14-16° (зимняя температура мелководных лагун Калифорнии).
- Глубина 3-40 м, донная температура 21,4 и 13,2°. Характерны многочисленные Nonion scapha (Fichtel et Moll), Elphidium, Buliminella elegantissima (Orbigny), Eponides ornatus (Orbigny) и др.
- Глубина 40-275 м, температура 13,2 и 8,5°. Встречаются Cassidulina caliphornica Cushman et Hughes, Eponides repandus (Fichtel et Moll), Polymorphina charlottensis Cushman, Quinqueloculina akneriana Orbigny, Robertina scharlottensis (Cushman), Sigmomorphina frondiculariformis (Galloway et Wissler), Triloculina trigonula (Lamarck). У побережья Центральной Америки - Buliminidae.
- Глубина 275-1800 м, температура 8,5 и 4°. Обнаружены Cassidulina cushmani Stewart et Stewart, Bolivina argentea Cushman, Bolivina subadvena var,spissa Cushman, Globobulimina pacifica Cushman, Uvigerina peregrina Cushman, Pseudoparrella pacifica (Cushman). Зона следует у берегов Центральной Америки за изотермой 8° (на 190 м глубже, чем у Калифорнии).
- Глубина свыше 1800 м, температура 4 и 2,4°. Bulimina rostrata Brady, Nonion pacificum Cushman (у побережья Центральной Америки соответственно с 4140 и 2500 м) и Pullenia bulloides (Orbigny). Весьма существенно, что Натлэнду удалось обнаружить сходные комплексы фораминифер в ископаемом состоянии в Калифорнии и выяснить, исходя из данных изучения современной зональности, условия осадкообразования в отдельные моменты неогеновой эпохи.
Приведенные данные вполне определенно указывают на зависимость батиметрического распределения фораминифер в условиях солености, в основном близкой к нормальной, от температуры. Фактор солености имеет несколько иное значение. Фораминиферы, как правило, стеногалинные организмы, способные существовать лишь в условиях, ограниченных пределами колебаний нормальной солености морской воды. Лишь некоторые виды способны выдерживать известное повышение солености (например обитающие в Красном море Orbitolites при солености до 43%о и др.). Условия более или менее значительного опреснения в прибрежных частях современных морей, в их заливах, во внутренних морях, эстуариях и реликтовых бассейнах выдерживают лишь немногие эвригалинные виды фораминифер (в основном из родов Proteonina, Аттобасишев, Trochammina, Streblus, Elphidium и др., а также некоторые Miliolidae); в условиях очень значительного опреснения встречаются некоторые приспособившиеся к этим условиям специфические формы, не свойственные бассейнам с нормальной соленостью.
Характеристику фораминифер ряда танатоценозов залива Аго (полуостров Сима, префектуры Миэ, Япония) дает Морисима (Morishima, 1948).
Глубина залива до 25 м, рН 8,10-8,40, температура 20,2-23,5°, соленость 32,1-33,3%o, т. е. лишь незначительно пониженная в сравнении с нормальной. У входа в залив танатоценоз пелагических, занесенных сюда из открытого моря форм: Globigerinidae, Globorotalia, Tretomphalus, далее в проливе, ведущем в залив Аго, танатоценоз Amphistegina radiata, Elphidium crispum (Linne), более обильный на песчаном, чем на скальном грунте, Rotalia rosea Orbigny,·Cibicides refulgens Montfort, Quinqueloculina bradyana Cushman. Центральная часть залива занята танатоценозом Rotalia papillosa Orbigny, Quinqueloculina lamarckiana Orbigny, Textularia аff. foliacea Heron-Allen et Earland; все эти формы наиболее обильны на илистом грунте. В многочисленных мелких заливах - ответвлениях залива Аго танатоценозы Trochammina globigeriniformis (Parker et Jones), Haplophragmoides, Elphidium fabum (Fichtel et Moll). У входа в ответвления залива к этому комплексу видов примешиваются формы смежного танатоценоза.
Несколько иные комплексы фораминифер приводятся Фледжером и Уольтоном (Phleger and Walton, 1950) для залива Марш (штат Массачусетс). Здесь выделяются следующие зоны, на составе комплексов фораминифер которых сказывается влияние солености.
- Зона водорослей Spartina patens и S. glabra (первая заливается кратковременно, вторая погружается при каждом приливе). Соленость во время прилива 31,5-32%o при отливе 25-31%o. Грунт, переполненный органическим веществом, алеврит или песок. Богатая фауна фораминифер: Trochammina inflata, Т. lobata, Т. cf. rotaliformis, Т. (Jadammina) macrescens, Мiliammina fusca. Эти виды встречаются и в других зонах, но здесь в наибольшем количестве. Специфические формы: Аrmorella sphaerica, Webbinella (?) sp. и Valvulineria sp. Виды Elphidium практически отсутствуют.
- Зона литоральных отмелей. Соленость 21-31%o грунт песок или алеврит с незначительным содержанием органики, с фрагментами раковин. Сильные приливно-отливные течения. Растительность отсутствует. Бедная фауна фораминифер: главным образом Trochammina squamata, реже Т. (Jadammina) macrescens.
- Зона зостеры всегда погруженная. Соленость в проливе 31-32%о, в самом заливе 27,5-31,5%o. Грунт алеврит, песок, вплоть до грубозернистого; органики мало. Trochammina squamata, Cribroelphidium incertum, Elphidiella arctica и формы первой зоны, но в иных процентных отношениях.
- Зона открытого залива Кейп Код. В мелководной (до 20 м) части грунт песчанистый с Eggerella advena и Proteonina atlantica, составляющими 90% микрофауны. В более глубоководной части (20-30 м) грунт илистый, либо песчаный, с более богатой фауной фораминифер: преобладающие Cribroelphidiumincertum, Elphidiella arctica и Trochammina squamata, кроме того, Ammodiscus sp., Ammotium cassis, Elphidium articulatum, Eponides frigidus var. calidus, Glomospira, cf. gordialis, Quinqueloculina seminulum и др. Reophax scorpiurus Montfort, R. scotti, R. curtus.
Для эстуарии реки Ранс (Франция) (Marie, 1938), где соленость с 32-36%o у Сен-Мало падает до 0,07%o в наиболее удаленных от моря частях эстуария, встречаются Elphidium cf. excavatum (Terquem), Astrononion cf. stelligerus (Orbigny), Lagena hexagona (Williamson), Тrochammina cf. nitida Brady, Spiroloculina aff. grata Terquem, Bolivina punctata Orbigny, Patellina corrugata Williamson, Globigerina bulloides Orbigny, Spirillina vivipara Ehrenberg и Streblus beccarii (Linne). К различным условиям солености приурочены те или иные из перечисленных видов.
Своеобразные комплексы фораминифер были описаны из залива Яде - устье реки Везера (Вагtenstein, 1938; Bartenstein и Brand, 1938). В различных частях залива в связи с неодинаковым его опреснением наблюдаются свои комплексы фораминифер. В подвергающейся частичному осушению при отливе области солоноватоводного мелкоморья ваттов первым из названных авторов определены: Proteonina difflugiformis (Brady), Р. fusiformis Williamson, Haplophragmoides canariensis (Orbigny), Ammobaculites agglutinans (Orbigny), Trochammina inflata (Montagu), Jadammina polystoma Bartenstein et Brand, Quinqueloculina (Мiliammina?) arenacea (Rhumbler), Miliammina fusca Brady. Наиболее характерна Jadammina polystoma (семейство Trochamminidae), обладающая хитиноидной раковиной, в состав которой включены отдельные песчинки. Помимо обычного для Trochammina базального устья на септальной поверхности последней камеры раковины расположено 3-7 дополнительных отверстий.
В качестве примера фауны фораминифер бассейнов с пониженной соленостью можно привести фауну современного Черного моря (средняя соленость - 19%o), откуда известно всего 12 видов фораминифер из родов Ammobaculites, Verneuilina, Quinqueloculina, Nonion, Elphidium, Discorbis. В области распространения илистых грунтов массового развития достигает Streblus beccarii (Linne) (Долгопольская и Паули, 1931).
Из Каспийского моря, отличающегося значительными колебаниями солености (12-13%o в среднем и до 50%o и более в некоторых его заливах) известно 5-6 видов. Наиболее распространенные из них - Streblus beccarii (Linne) и Elphidium poeanum (Orbigny) - являются в широком смысле эвригалинными; они обнаружены в условиях солености от 7,38%o до 50,0%o. Отмечается, что у Streblus beccarii в условиях пониженной солености раковины становятся более тонкими, почти лишенными скульптуры; пупок делается зияющим из-за исчезновения пупочной втулки. Наоборот, при значительном повышении солености (45-50%o) пупок заполнен известью. Раковины Streblus в условиях слабого опреснения и нормальной солености (25-35%o) наиболее крупные; они сильно мельчают при увеличении солености до 40%o и выше (данные В. А. Шохиной).
Влияние опреснения сказывается на раковинах Streblus beccarii в том же смысле и в Немецкой бухте Северного моря; при значительном опреснении раковины этого вида становятся здесь почти чисто псевдохитиновыми (Pratje, 1931).
В Аральском море (средняя соленость 11%o) обитают всего два вида фораминифер из тех же родов. Необходимо отметить, что фауна фораминифер современного Средиземного моря (соленость 37-39%o) насчитывает свыше сотни видов и имеет типично морской характер.
Сравнение основных количественных признаков средиземноморских (С) и черноморских (Ч) Streblus beccarii (Linne)
(по Ливеровской из Раузер-Черноусовой и Фурсенко. 1937)| Признаки | Бассейн | Число измерений | Средняя величина (М) |
| Диаметр раковины | С Ч | 50 51 | 1,035±0,011 мм 0,831±0,022 мм |
| Число камер | С Ч | 50 53 | 28,00±0,01 кам. 20,06±0,37 кам. |
| Число оборотов спирали | С Ч | 50 54 | 2,62±0,04 об. 2,40±0,05 об |
| Диаметр начальной камеры | С Ч | 47 52 | 0,080±0,003 мм 116±0,003 мм |
Весьма показательно соотношение количественных признаков у Streblus beccarii из Адриатического моря (окрестности Венеции) и Черного моря (район Карадага). Как видно из нижеприведенной таблицы, черноморская форма мельче; ее раковина образована меньшим числом камер при почти том же числе оборотов. Диаметр начальной камеры у черноморских Streblus больше, но не настолько, чтобы можно было отнести их к микросферической генерации (в расчет при измерениях принимались только мегасферические особи, однако без выделения генераций А1 и А2). Средиземноморская форма богато скульптурирована и имеет пупочную втулку, а черноморская лишена скульптуры, и пупок у нее зияющий.
Не исключена возможность, что черноморские Streblus beccarii принадлежат особому подвиду, признаки которого выработались под влиянием опреснения и в какой-то мере наследственно закрепились.
Влияние периодического понижения солености на рост раковины хорошо видно на примере Elphidium striato-punctatum (Fichtel et Moll), существующего в эстуарии Эльбы - Куксгафене в зоне, где заметно снижается влияние приливов (соленость 20%o) и отливов (17%o). Неравномерность возрастания камер зависит от того, на какой момент падает возникновение той или иной из них. Это явление Румблер (Rhumbler, 1911 -1913) объясняет изменением осмотического давления в условиях различной солености (рис).
Особый интерес представляют реликтовые фауны фораминифер, сохранившиеся в бассейнах, давно потерявших связь с морем. Миоценовые реликты известны из соленых озер Семиградья (Венгрия), откуда Дадаем (1884) была описана Entzia tetrastomella Daday - форма с псевдохитиновой раковиной, агглютинирующая кремневые пластинки. Entzia представляет собою, по-видимому, конечное звено в ряду Trochammina - Jadammina.
В солоноватоводных реках, впадающих в озеро Эльтон (Нижнее Заволжье) В. Крапин (1929) обнаружил раковины Trochammina sp. и Cyclammina (?А. Ф.), возможно, ископаемые, вымытые из третичных пород. Из самосадочного реликтового озера Балпаш-сор (Кокчетавская область) была описана Borovina zernovi Schmalhаusеn (Шмальгаузен, 1951). Раковина у Borovina, по данным Шмальгаузен, хитиноидная, с внедренными кремневыми пластинками. Наличие в стенке Borovina пор, а также то, что септы у нее двойные, говорит о том, что этот род вряд ли близок Entzia; скорее он является родственным Streblus, представители которого широко распространены в Понто-Арало- Каспийском бассейне. В озере Балпаш-сор были найдены лишь раковины отмерших Borovina; предполагается, что Borovina обитают в грунтовых водах, питающих озеро; попадая в это последнее они гибнут из-за слишком высокой минерализации (до 150,8%o).
Наконец, реликтовые же, по-видимому, формы были найдены А. Л. Бродским (1928; см. также у Зернова, 1949) в колодцах пустыни Каракум, вскрывающих воды грунтового бассейна площадью 5000 км2 и глубиной до 20 м; соленость 5-11%o, температура 17-20°, аэрация слабая. Фораминиферы, по мнению Бродского, обитают в капиллярных пространствах, поперечник которых всего лишь в 10-15 раз превышает диаметр раковин. Фораминиферы грунтовых вод Каракумов небольших размеров (не свыше 0,16 мм), имеют хитиноидную, почти лишенную извести, округло-удлиненного очертания раковину и относятся к установленным Бродским видам родов Spiroloculina, Pyrgo, Textularia (?). Кроме того А. Л. Бродским указываются Lagena, Nodosaria, Globigerina и Discorbis. Представители последнего рода могут быть видоизмененными Streblus beccarii, что же касается остальных трех, то правильность определения их сомнительна. По мнению Брод-ского (1928), изученные им фораминиферы реликты миоценового средиземноморского бассейна, заходившего на территорию Каракумов. Д. Ф. Николюк (1948) обнаружил в ряде колодцев Каракумов фауну фораминифер, сходную с описанной Бродским. Отмечается, что наиболее благоприятная для существования фораминифер соленость, близка к нормальной океанической (рис).
Фораминиферы обитают между песчинками грунта в заполненных водой капиллярных пространствах с поперечником 0,25-1,5 мм. Фораминиферы каракумских грунтовых вод рассматриваются как реликты фауны акчагыльского бассейна. Аналогичные находки известны из солоноватоводных и почти пресных грунтовых бассейнов северо-запада Сахары (Уэд Рир, к югу от Бискры), откуда Готье-Льевром (Gauthier-Lievre, 1955) были определены Ammodiscus, Miliolidae, Ophthalmidium, Trochammina, Nonion, Аnоmalina и Cibicides.
Из прочих факторов на развитие фораминифер влияет свет, в основном благодаря жизнедеятельности водорослей симбионтов - зооксантел, обитающих в протоплазме Amphistegina, Peneroplis, Sorites, Marginopora и др. Большое значение имеет пища, количество которой сказывается в первую очередь на плотности населения фораминифер. Шохина отмечает, что обилие экземпляров Streblus beccarii (Linne) и Elphidium poeanum (Orbigny) стоит в прямой зависимости от развития водорослей и служащих фораминиферам пищей аэробных бактерий. Наоборот, при отсутствии водорослей в местах, где изобилуют гнилостные бактерии, названные виды фораминифер хотя и встречаются, но в значительно меньших количествах. Здесь, по-видимому, существенную роль играет и газовый режим. При недостатке пищи, в частности диатомовых водорослей, могут развиваться угнетенные фауны карликовых фораминифер, как это имеет место в области современного Патагонского шельфа (Boltovskoy, 1954); при этом некоторые виды утрачивают присущую им скульптуру, раковины других становятся асимметричными. Большинство фораминифер обитает в условиях насыщения воды кислородом или, во всяком случае, достаточного его содержания. Лишь некоторые солоноватоводные и морские формы из родов Haplopragmoides, Trochammina и др. могут развиваться при слабой аэрации. Это относится к чисто морским Cyclammina и Bathysiphon. Наличие карбоната кальция необходимо для существования форм с известковой раковиной. В теплых водах, где условия для биохимического выделения карбоната кальция наиболее благоприятны, развиваются фораминиферы с крупными, массивными известковыми раковинами.
Благодаря относительной однородности температурных условий, не только зональные, но и биогеографические отличия в составе фауны холодноводных бассейнов выражены значительно слабее, чем в субтропических и тропических областях. Практически, например, можно говорить о биполярном распространении холодноводных мелководных фораминифер (рис). Но все же для относительно однотипной арктической мелководной фауны могут быть установлены более или менее хорошо ограниченные биогеографические провинции (Щедрина, 1947, 1955; Саидова, 1955 и др.), Так, например, в арктических морях (бассейн Северного Ледовитого океана), помимо видов, встречающихся и в Тихоокеанском бассейне, обитают некоторые специфические формы. Для литоральной зоны это беломорская Trochammina cyclostomata Stschedrina, обладающая уплощенной раковиной с гибкой хитиноидной стенкой, благодаря чему способна менять свою конфигурацию, приспосабливаясь к узким пространствам, и «прилипать» к субстрату при отливе. Эндемичный для арктической сублиторали вид Elphidium gorbunovi Stschedrina. В псевдоабиссали встречаются Alveolophragmium orbiculatum var. caraensis Stschedrina и др.; там, где сказывается влияние атлантических вод, обнаружены Anomalina baltica (Schroeter) и А. coronata (Parker et Jones). Для верхней части батиальной зоны арктических морей характерен Cornuspiroides striolatus (Brady), для нижней - С. profundus Stschedrina. Таким образом, по данным Щедриной, фауны фораминифер дальневосточных и арктических морей, входящих в состав единой Арктической провинции в понимании Кешмэна. отличаются достаточно резко выраженными биогеографическими особенностями, проявляющимися, что необходимо особо подчеркнуть, в видовом составе.
Биогеографические отличия в составе фауны фораминифер сказываются в мелководных зонах теплых морей и в умеренном поясе в значительно более резкой степени, чем в холодноводных бассейнах. Кешмэн (Cushman, 1928) выделяет четыре основные биогеографические провинции распространения современных тепловодных фораминифер (рис). Средиземноморская провинция охватывает область западного побережья Африки и юга Европы, а также Средиземное и Красное моря и северо-западная часть Индийского океана. Вест-Индская провинция включает западную часть Атлантнки от южной границы Бразилии на юге до Бермудских островов на севере; отдельные, характерные для этой провинции формы распространены через весь Атлантический океан до берегов Ирландии. Восточно-Африканская провинция обнимает область восточного побережья Африки и значительную часть Индийского океана, в том числе западное побережье полуострова Индостана. Наконец, Индо- Тихоокеанская провинция ограничена пределами Бенгальского залива, восточной части Индийского океана, областью Индонезии и восточной части Тихого океана до берегов Японии на севере и до островов Гуамоту на востоке. Строго говоря, Кешмэн принимает Индо-Вест-Пацифическую провинцию в смысле Экмана (Ekman, 1935); Кешмэн подразделяет ее на две подпровинции: В осточ ноАфриканскую и Индо- Тихоокеанскую.
В свете более новых данных, фауна фораминифер, близкая, но не тождественная известной из северной части Индо-Тихоокеанской провинции, распространена в области западного побережья Северной Америки (Калифорния). Поэтому целесообразно установить еще одну биогеографическую область распространения тепловодных фораминифер - Воcточно-Тихоокеанскую.
В геологическом прошлом биогеографические особенности фаун фораминифер появляются впервые в девоне в виде обособления фауны уральского и платформенного типов (Е. В. Быкова, 1952). В верхнем палеозое проявляются резкие биогеографические отличия фаун фораминифер: развитие штаффелловых фаций на Русской платформе, в отличие от фаций с крупными псевдсфузулинами и парафузулинами в нижней перми. Еще более резко выражены биогеографические отличия в развитии лягенидовых фаций в эпиконтинентальных морях верхней перми - в цехштейне Прибалтики, в казанском ярусе Русской платформы, на севере Азиатской части СССР, в Северной Америке (Техас) и др., и параллельно развитие фузулиновых фаций (с Neoschwagerinidae и др.) в геосинклинальной области Тетиса и на Дальнем Востоке СССР, а также в Японии. Относительно нечетко выраженные биогеографические особенности фаун фораминифер отмечались для юрского периода и для нижнемеловой эпохи. Они становятся вновь более резко выраженными в верхнем мелу, в палеогене и неогене (см. «Историческое развитие фаун фораминифер»).
Попадая в осадок, раковины фораминифер привносят в него вещества, являющиеся продуктом их жизнедеятельности: углекислый кальций, отчасти - магний, окись железа и др. Деятельность фораминифер с точки зрения их биогеохимических функций определяется, помимо общих для всех организмов газовой и метаболической функций, функциями кальцитовой и концентрационной в отношении, главным образом, железа (Вернадский, 1940).
В образовании современных тепловодных осадков неритовой области бентические фораминиферы играют обычно относительно небольшую роль, образуя лишь более или менее заметную примесь к основной части осадка, терригенной или подчас карбонатной, органогенной или хемогенной (например, в коралловых песках и т. п.). Лишь в относительно редких случаях, в рифогенных фациях современных морей фораминиферы выступают в качестве породообразующих организмов наравне с кораллами, известковыми водорослями и другими организмами.
Прибрежные пески рифов Палау (Каролинские острова) состоят в основном из раковин, обитающих по соседству на водяных растениях фораминифер. У острова Сент-Винсент (Карибское море) Amphistegina lessoni образует на глубинах 12-91 м до двух третей осадка. На побережье Австралии и прилегающих к нему рифах прибрежные пески часто состоят из одних лишь раковин Orbitolites. Меньшее значение имеют донные фораминиферы как породообразующие организмы в осадках батиальной области, в терригенных илах которой они, однако, почти всегда встречаются в достаточном количестве. В области Гольфстрима донные агглютинированные фораминиферы образуют до 18% осадка (Wa1ther, 1893-1894).
В геологическом прошлом фораминиферы мелководных теплых областей неоднократно выступали в роли породообразующих организмов. В верхнем палеозое - при отложении карбонатных толщ нижнего и среднего карбона на Русской платформе, отчасти - на Урале; в верхнем карбоне и нижней перми - отчасти на Русской платформе(верхний карбон Самарской луки и др.) и на Урале (рис). В области западного склона среднего и южного Урала распространены крупные рифовые массивы, сложенные фузулиновыми известняками или смешанными органогенными - мшанковыми, криноидными, а также смешанного состава известняками, содержащими более или менее значительную примесь скелетов фораминифер. Породообразующими являются подчас донные фораминиферы в верхнепермских известняках и некоторых других геосинклинальных областей, а также в верхнемеловых орбитоидных и других фораминиферовых известняках Средиземноморской области, (рис) и, кроме того, наряду с планктонными формами, в белом мелу. В качестве породообразующих организмов бентические фораминиферы выступают также в нуммулитовых и дискоциклиновых известняках палеогена и в ряде областей Альпийской складчатости, в том числе на Кавказе, а также в Крыму и т. д. (рис). В неогене фораминиферы редко являются породообразующими, но, как и в более древних карбонатно-глинистых отложениях в составе последних раковины фораминифер имеют подчас довольно существенное значение. В массовых количествах в некоторых сарматских известняках Молдавии встречаются Nubecularia novorossica Karrer et Sinzov, почти нацело слагающие породу.
| "ЭБНБ" >> "Ф" >> "ФО" |
Статья про "Фораминиферы, экология и тафономия" в Энциклопедии БНБ была прочитана 5995 раз
| Коптим скумбрию в коробке |
| Ананасы на гриле |
TOP 15
- Вши и борьба с ними
- Пижма обыкновенная
- Каштан конский обыкновенный
- Роль элементов в жизни растений
- Пшеница
- Иван-чай узколистный
- Обеззараживание химическими средствами
- Календула лекарственная
- Клевер луговой
- Дезинфекция текущая
- Душица обыкновенная
- Бессмертник песчаный
- Как устроены растения
- Морфологические признаки почвы
- Хвощ полевой