
БНБ
"ЭБНБ" (14252)
- "Я" (73)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Ядерные рецепторы
Определение "Ядерные рецепторы" в ЭБНБ
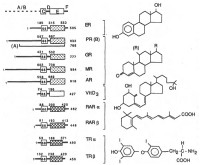 |
| Ядерные рецепторы и их лиганды |
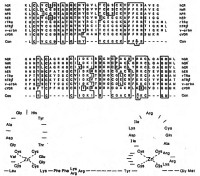 |
| Цинковый палец |
 |
| Аминокислотные последовательности ядерных рецепторов |
При воздействии на клетки-мишени стероидным гормоном (например, кортизолом) в условиях in vivo или in vitro образуется два различных комплекса стероид-рецептор. Незначительная фракция меченного гормона обнаруживается в супернатанте или цитозольной фракции гомогената тканей, тогда как большая часть инкорпорированного стероида связана с белком в ядерной фракции и освобождается лишь при дополнительном экстрагирующем воздействии. Показательно, что ассоциация гормон-рецепторного комплекса с компонентами ядерной фракции (т.е. ДНК) прочнее, чем свободного рецептора.
Существенное отличие в связывании тиреоидных гормонов и ретиноевой кислоты состоит в распределении гормон-рецепторного комплекса лишь в ядерной фракции, что подробно рассматривается далее.
Последующими исследованиями было достоверно доказано, что индуцируемая лигандом/гормоном трансформация нативного рецептора присуща рецепторам всех классов стероидных гормонов, но не тиреоидным гормонам. Трансформация рецептора эстрогенов сопровождается также увеличением константы седиментации, что связано с димеризацией рецептора. Хотя трансформация рецептора не обязательно сопровождается его спонтанной димеризацией, установлено, что рецепторы прогестагенов, глюкокортикоидов и тиреоидных гормонов взаимодействуют с чувствительными последовательностями ДНК ядра в димеризованной форме. Оказалось также, что трансформация и димеризация рецепторов стероидных гормонов нередко может протекать с большей скоростью через стадию уменьшения Ks. Этот феномен получил объяснение после описания взаимодействия нативного рецептора с белками теплового шока - разновидностью клеточных chaperons (белков, которые способствуют стабилизации 2-ной структуры мРНК и ряда белков и синтезируются в ответ на предъявление клетке чрезвычайных стимулов). Возможно, что chaperon'bi занимают участок связывания рецептора с чувствительной нуклеотидной последовательностью и освобождают его при трансформации в необычных внутриклеточных условиях (например, в присутствии эндогенных анионов металлов или аминофосфоглицерола). Освобождение места связывания наступает при реакции стероида с анионным центром или в результате индукции аутопротеолитической активности в белках теплового шока. Точное значение этих взаимодействий остается пока неизвестным, хотя можно предполагать их усиливающую и/или разрешающую роль.
Ряд исследований показал, что рецепторы тиреоидных гормонов после их синтеза подвергаются транслокации в ядро и перманентно существует в ДНК-связанной форме (без связи с белками теплового шока) и, таким образом, не претерпевают лиганд-индуцированной трансформации. Также не обнаружено цитозольной формы рецепторов тиреоидных гормонов и ретиноевой кислоты. Клетки чувствительных к действию ретиноевой кислоты тканей содержат так называемый клеточный связывающий белок, CRABP, который может служить цитозольным переносчиком гормона.
Современное прочтение теории двухстадийной активации рецепторов стероидных гормонов заключается в дискуссионности вопроса о присутствии цитозольного рецептора и необходимости его трансформации и транслокации в ядро. Большинство исследователей разделяет мнение о валидности двухстадийной активации рецептора. Спорным остается положение о цитоплазменно-ядерном равновесии распределения рецепторов сам механизм транслокации.
| "ЭБНБ" >> "Я" |
Статья про "Ядерные рецепторы" в Энциклопедии БНБ была прочитана 8369 раз
| Бургер двойного помола |
| Английская картошка фри |
TOP 15
- Вши и борьба с ними
- Пижма обыкновенная
- Каштан конский обыкновенный
- Роль элементов в жизни растений
- Пшеница
- Иван-чай узколистный
- Обеззараживание химическими средствами
- Календула лекарственная
- Клевер луговой
- Дезинфекция текущая
- Душица обыкновенная
- Бессмертник песчаный
- Как устроены растения
- Морфологические признаки почвы
- Хвощ полевой