
БНБ
"ЭБНБ" (14252)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Тиреоидные гормоны, дейодиназная регуляция метаболизма
Определение "Тиреоидные гормоны, дейодиназная регуляция метаболизма" в ЭБНБ
 |
| Метаболизм тироксина |
 |
| Селеноцистеин и цистеин |
- примерно 1/3 общего пула Т4 подвергается монодейодированию в периферических тканях с образованием Т3, который обладает наибольшей биологической активностью;
- около 1/2 Т4 монодейодируется в периферических тканях с образованием рТ3, который не обладает калоригенной активностью;
- большая часть оставшейся 1/6 от общего количества тироксина экскретируется с желчью и мочой в виде конъюгатов с серной и глюкуроновой кислотами;
- небольшая часть (менее 5%) превращается в тетрайодтироуксусную кислоту путем окисления аланиновой боковой цепи (дезаминирование и декарбоксилирование), а также путем разрыва диэфирной связи.
Уровень T4S может составлять в сыворотке пуповинной крови новорожденного до 21 нг/л по сравнению с 1,6 нг/л в сыворотке крови взрослого. Считается, что по крайней мере 20% образующегося за день Т3 метаболизируется в реакциях сульфоконъюгации.
Дейодиназы имеют особую биологическую значимость в последовательной деградации молекулы Т4 как через каскадный путь образования его производных Т3, рТ3, различных дийодтиронинов (Т2), монойодтиронинов (Т,) и тиронина (Т0), так, по-видимому, и в разрушении конъюгатов тиреоидных гормонов с остатками серной и глюкуроновой кислот, также как и в разрушении продуктов окислительного дезаминирования и декарбоксилирования тиреоидных гормонов.
5'-дейодиназа 1-го типа (5'-ДI)
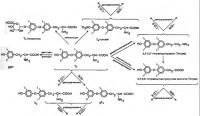
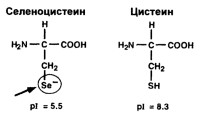
Участвует в превращении Т4 в Т3 и рТ3 в 3,3'-Т2, Т3 в 3,5-Т2 и далее части дийодтиронинов в монойодтиронины (Т1) и последних в свободный от йода тиронин (Т0) (см. рис.). Глюкуроновые и сульфоконъюгаты Т4 и Т3 также дейодируются при участии 5'-ДI. Активность 5'-ДI полностью блокирутся пропилтиоурацилом и ауротиоглюкозой.5'-ДI представляет собой мембраносвязанный полипептидный фермент, активность которого уменьшается вне липидного окружения (например, при делипидировании - удалении фосфатидилхолина и фосфатидилэтаноламина с помощью детергентов). Активность 5'-ДI зависит от температуры и рН среды. Наибольшая активность проявляется при рН 6,5-7,5. При сравнении рН оптимума для проявления активности 5'-ДI и 5-ДIII при дейодировании различных йодтиронинов, было обнаружено, что в зависимости от величины рН изменяется конформация тех или иных активных центров этих ферментов. Возможно, что эти изменения рН являются определяющими в том, какое кольцо будет дейодировано. Так, например, активность 5-ДIII повышается при рН>8,0. Реакции дейодирования, осуществляемые 5'-ДI зависимы от присутствия и ускоряются аминокислотой селеноцистеином. Селеноцистеин, по- видимому, участвует в формировании активного центра фермента.
Недостаточное поступление селена в организм с водой и пищей приводит к нарушению превращения Т4 в Т3, а в условиях йодной недостаточности дополнительное поступление селена в организм, сопровождаемое повышением активности 5'-ДI, ведет к ускорению метаболизма Т4 и ухудшению течения гипотиреоидизма, так как щитовидная железа в условиях йодного дефицита не может компенсировать ускоренный метаболизм этого гормона.
Наибольшее содержание 5'-ДI обнаружено в печени, почках, и более низкое - в щитовидной железе, скелетных и сердечной мышцах и других тканях. 5'-ДI определяет эффективность воздействия тиреоидных гормонов на клетку, так как от ее активности зависит какое
количество Т4 превратится в его более активный метаболит
Проявление многих эффектов тиреоидных гормонов при гипертиреоидизме обусловлено отчасти повышением активности 5'-ДI и обеспечиваемым этим более высоким уровнем Т3.
Дейодиназы локализуются в различных субклеточных фракциях, например, в печени - в мембранах эндоплазматического ретикулума гепатоцитов. Не обнаружено активности 5'-ДI в цитозоле, митохондриях, лизосомах или ядрах гепатоцитов. Фермент отсутствует в купферовских и эндотелиальных клетках.
В почках активность 5'-ДI зарегистрирована в плазматических мембранах и эндоплазматическом ретикулуме эпителиальных клеток проксимальных канальцев. Эти клетки, в частности, эффективно захватывают профильтровавшийся рТ3, который реабсорбируется, подвергается дейодированию и поэтому отсутствует в моче, в то же время 3,3-Т2, образовавшийся из рТ3, присутствует в моче.
Активность 5'-ДI, как и 5'-ДII, обнаружена в тяжелых фракциях синаптосомальных мембран и мембран нервных терминалей. Активность коррелирует с содержанием в этих фракциях Na/K+-АТФазы, Т4 и Т3. Активность 5-ДIII обнаруживается в легких фракциях мембран нервных окончаний и ликворе.
Многие соматические заболевания, такие как цирроз печени, прогрессирующие злокачественные опухоли, почечная недостаточность, инфаркт миокарда, лихорадочные состояния, ожоги, травмы и др., сопровождаются подавлением активности 5'-ДI, осуществляющей превращение Т4 в Т3 и рТ3 в 3,3'-Т2. Подобное действие могут оказывать на активность 5'-ДI некоторые лекарственные средства. Например, активность фермента подавляется пропилтиоурацилом, который оказывает положительный терапевтический эффект при гипертиреоидизме. Результатом понижения активности 5'-ДI является понижение уровня Т3 в крови и повышение уровня рТ3. Амиодарон и йоподат также нарушают внутриклеточное превращение Т4 в Т3.
Из фактов, свидетельствующих о зависимости уровней Т3 и рТ3 в крови и тканях от активности дейодиназ следует, что, основываясь только на данных о понижении сывороточных уровней Т3 и повышении рТ3, нелегко решить, являются ли эти изменения уровней гормонов отражением нарушения функции щитовидной железы или это результат нарушения метаболизма тиреоидных гормонов, обусловленный соматическими заболеваниями и (или) лекарственной терапией. Выздоровление или отмена лекарственных средств могут приводить к восстановлению уровней Т3 и рТ3 в крови и облегчению дифференциальной диагностики.
5'-дейодиназа II-го типа (5'-ДII)
Этот фермент был открыт в 80-е годы при анализе механизмов образования внутриклеточного Т3 в условиях блокады активности 5'-ДI пропилтиоурацилом. 5'-ДII также, как и 5'-ДI, является мембраносвязанным ферментом. Для проявления своей активности требует присутствия восстановленных тиольных соединений, которые должны связаться с активными центрами фермента одновременно с йодтиронинами (Т4, рТ3) и только после этого протекание реакции дейодирования становится возможным. На рис. (сплошными стрелками) показано, какие субстраты подвергаются дейодированию при воздействии 5'-дейодиназ I и II типов.В плаценте, бурой жировой ткани, коре мозга, миокарде, скелетных мышцах и самой щитовидной железе блокада 5'-ДI дейодиназной активности пропилтиоурацилом не полностью устраняет превращение Т4 в Т3. Это превращение осуществляется под действием фермента 5'- ДII дейодиназы, активность которой не блокируется пропилтиоурацилом.
5'-ДII дейодиназа отличается от 5'-ДI как характером распределения в тканях, так и активностью в процессе развития тканей в онтогенезе организма и в условиях патологии. В ЦНС ее активность наиболее высока в плазматических мембранах нервных терминалей и дендритов, глиальных клеток, а в переднем гипофизе - в мембранах эндоплазматического ретикулума. 5'-ДII активность проявляется в 2-х типах клеток аденогипофиза - тиротрофах и соматотрофах, в которых имеет место действие тиреоидных гормонов. Это подтверждает важную роль 5'-ДII в центральной нервной системе. Вероятно, роль 5'-ДII в мозге заключается в поддержании в нем на постоянном уровне содержания Т3. Установлено, что понижение уровня Т4 в крови ведет к быстрому увеличению содержания и активности 5'-ДII в мозге и, в частности, в клетках гипоталамуса и гипофиза. Наоборот, при повышении уровня Т4 в крови, понижается активность 5'-ДII в мозге. Содержание 5'-ДII в мозге зависит также от уровня других продуктов метаболизма Т4.
Таким образом, благодаря наличию зависимости активности нейрональной дейодиназы II-го типа, от уровня Т4 в крови, мозг не только способен поддерживать на постоянном уровне содержание Т3, но и способен защищать клетки от воздействия избыточного уровня Т3.
По крайней мере у эутиреоидных крыс, 5'-ДII обеспечивает образование около 50% Т3 из Т4. Т4 является для дейодиназы 2-го типа у этих животных более предпочтительным субстратом в сравнении с 5'- Д!, которая катализирует реакции деградации рТ3. Около 90% Т3 образуется в мозге и мышцах у гипотиреоидных крыс при участии 5'-ДII. Поглощение этими тканями Т4 увеличивается при гипотиреоидизме на 30-50%, что способствует увеличению образования в них Т3.
Обычно активность 5'-ДII ниже активности 5'-ДI и, как полагали, ее главная роль заключается в образовании Т3 из Т4 внутри клеток для локального обеспечения потребностей прежде всего таких органов, как мозг, гипофиз, плацента и бурая жировая ткань. Однако, следующие факты свидетельствуют о том, что дейодиназа II-го типа играет важную роль в образовании Т3, циркулирующего в крови.
5'-ДII дейодиназа обеспечивает в нормальных условиях превращение в щитовидной железе в трийодтиронин около 29% тироксина, а превращение остального количества Т4 обеспечивается при участии 5'-ДI. В условиях стимуляции функции щитовидной железы уровень мРНК этого фермента резко возрастает, повышается активность 5'-ДII. Вероятно, что главным фактором, определяющим активность дейодиназы ll-типа, является концентрация свободного Т4 в тироцитах щитовидной железы. Повышение образования Т4 в щитовидной железе при ее активации ведет к тому, что при участии дейодиназы II-типа образуется около 60-80% Т3 из Т4.
5'-ДII обспечивает относительно высокий уровень образования Т3 и повышение отношения Т3/Т4 при заболеваниях щитовидной железы, сопровождающихся повышением ее функциональной активности, в частности, при болезни Грейвса. Практическое значение имеет и тот факт, что активность 5'-ДII не ингибируется у пациентов при применении пропилтиоурацила.
5-дейодиназа Ill-го типа (5-ДIII)
Этот фермент участвует в дейодировании тирозильного, внутреннего кольца йодтиронинов, удаляя атомы йода в положениях 3 и 5. Фермент имеет важное значение в метаболизме Т3, посредством превращения его в 3,3'-дийодтиронин, и в реакциях инактивации Т4, посредством превращения тироксина в неактивный рТ3 (рис.). Этот фермент активен во многих тканях, особенно таких как печень, плацента, кожа, мозг. В передней доле гипофиза активность фермента пока не обнаружена. Известно, по крайней, мере две изоферментные формы 5-ДIII, которые как и 5'-ДI, являются селеноцистеинзависимыми.Дейодирование внутреннего кольца является ключевым путем метаболизма тиреоидных гормонов, который ведет к инактивации метаболитов. Этот эффект достигается действием 5-ДIII, активность которой зависит от функционального состояния щитовидной железы и изменяется в процессе онтогенетического развития. Кроме того, установлено, что Т3 и ретиноевая кислота также влияют на активность 5-ДIII и уровень ее мРНК в культуре адипоцитов бурой жировой ткани.
Высокая активность 5-ДIII обнаружена в эндоплазматическом ретикулуме гепатоцитов. Предполагается, что 5-ДIII активируется тем же путем, что и 5'-ДI. Эти две дейодиназы располагаются в одинаковых субклеточных фракциях и тесно структурно связаны друг с другом. Активность этого селеноцистеинзависимого фермента также угнетается пропилтиоурацилом и ауротиоглюкозой, хотя для достижения ингибирующего эффекта требуется 10-кратная концентрация этих агентов в сопоставлении с таковой для дейодиназы I типа. Активность 5-ДIII зависит от присутствия восстановленных тиолов. Оптимум рН для 5-ДIII равен 8-8,5, что позволяет ей осуществлять избирательный катализ дейодирования внутреннего кольца и других субстратов, нечувствительных к действию 5'-ДI, 5-ДIII активно дейодирует сульфоконъюгаты йодтиронинов, что также способствует инактивации Т4.
5-ДIII и ее изоэнзимы нестабильны, что затрудняет изучение их свойств. Трудно установить точное место их локализации в тканях, но вероятно, в ЦНС фермент связан с микросомальными мембранами астроглии, а в плаценте с мембранами клеток хориона. Уровень 5-ДIII повышен при гипертиреоидизме и понижен при гипотиреоидизме и полагают, что дейодиназа III-го типа, подобно дейодиназе II-го типа, может защищать мозг и другие ткани плода от избыточного уровня или недостатка Т4.
Обобщающая характеристика участия различных типов дейодиназ в регуляции метаболических превращений тиреоидных гормонов
| 5'-дейодиназа I типа | 5'-дейодиназа II типа | 5-дейодиназа III типа | |
| Принадлежность отщепляемых атомов йода | фенольный и тирозильный радикалы (внутреннее и внешнее кольца) | фенольный радикал (внешнее кольцо) | фенольный радикал (внешнее кольцо) |
| Предпочитаемый субстрат | рТ3>>Т4>Т3 | Т4>рТ3 | Т3>Т4 |
| Константа Михаэлиса (Km) для тироксина | ~1 мкмоль | ~1 нмоль | ~40 нмоль |
| Тканевое распределение | печень, почки, щитовидная железа | ЦНС, бурая жировая ткань, гипофиз | ЦНС, плацента, печень, кожа |
| Кинетический механизм | пинг-понг | последовательныйпоследовательный | |
| Влияние пропилтиоурацила | угнетает | не влияет | не влияет (?) |
| --//-- йодоацетата | сильно угнетает | слабо угнетает | угнетает |
| --//-- йопановой кислоты | угнетает | угнетает | угнетает |
Сокращения:
рТ3 - 3,3',5'-трийодтиронин, реверсивный трийодтиронин
Т0 - тиронин
Т2 - дийодтиронин
Т3 - 3,5,3'-трийодтиронин
Т4 - 3,5,3',5'-тетрайодтиронин, тироксин
| "ЭБНБ" >> "Т" >> "ТИ" |
Статья про "Тиреоидные гормоны, дейодиназная регуляция метаболизма" в Энциклопедии БНБ была прочитана 11498 раз
| Коптим скумбрию в коробке |
| Гороховое пюре |
TOP 15
- Вши и борьба с ними
- Пижма обыкновенная
- Каштан конский обыкновенный
- Роль элементов в жизни растений
- Пшеница
- Иван-чай узколистный
- Календула лекарственная
- Обеззараживание химическими средствами
- Клевер луговой
- Дезинфекция текущая
- Душица обыкновенная
- Бессмертник песчаный
- Как устроены растения
- Морфологические признаки почвы
- Хвощ полевой